�t�ɐ�֑k��܂ł̐��Ԃɂ��Đ������܂��B�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�쐬�F2003/12/15
�����ł͛z�������A���̎e�����C�܂ŗ������A�t���܂Ő������@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@![]() �@�@
�@�@![]()
�t�ɐ�֑k��܂ł̐��Ԃɂ��Đ������܂��B�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�@�쐬�F2003/12/15
�X�V�F�@04.05.09�@�i�P�j�C�ł̉�V�o�H�ɁA![]() �����V���L���i04.05.07�j��lj��B
�����V���L���i04.05.07�j��lj��B
05.03.15�@�P�D�͌�����C�ւɁA�u�O�͘p���ŃA���e�t���̐����v��lj�
�u����͌����ӊC��ɂ�����A���̐��ԁv�Ƀ����N�B
�@�k�O�u���l
�@���⓮���̑̉t�i���t�j�̉����Z�x�́A���Â̐̂ɁA�����C�����ցA�������痤��ցA�܂��삩��C���ڂ�Z�����̌��n�̊C�̉����Z�x�����݂��p����Ă���̂��Ƃ����Ă���B
�@�̉t�̉����Z�x�́A�W�����͖�O�D�U���A�l���܂ߗ��㓮���͖�O�D�X���A�C�����͖�P�D�P���ł���B
�@�i���݂̊C�̉����Z�x����3.5���ŁA�C�̖��Ғœ����̑̉t�͊C�̉����Z�x�Ɠ����ł��B�j
�@�ɂ̃^�b�v����������w�����̔����Ƒ̉t�̐Z���������x���ǂ����B
�@�삩��C�ցA���̋t�ɊC���������ړ����邱�Ƃ͒ʏ�͂ł��Ȃ��B����́A�̉t�i���t�j�̉����Z�x�����ɕۂ��߂̎d�g�݂��A�W�����ƊC�����ł͑S���t�̎d�g�݂ɂȂ��Ă��邩��ł���B
�W�����ƊC�����ł́A�̉t�̉������Z�x�����ɕۂ����݁i�Z�����̒����@�\�j���S���t
�@�W�����ł́A�҂�畆���琅���̓��ɟ��݂��݁A�������Ȃ���ΐ��Ԃ���ɂȂ�זE���j�Ă��܂��B����A�C�����ł͑̓��̐������C�����ɟ��ݏo���čזE�����ڂ�ł��܂��B
�@���������āA�W�����͑̂ɟ��ݍ���ł��鐅��̊O�ɏo���Ȃ���Ȃ�Ȃ����A�C�����͑̂���o�Ă��������ǂ�ǂ�⋋���Ȃ���A�����Ă����Ȃ��B���ꂪ�A�̉t�̉������Z�x�����ɕۂ��Ȃ���Ȃ�Ȃ����R�ł���B
�@�̉t�̉����Z�x��W�����͂O�D�U���A�C�����͂P�D�P���ɕۂ��߂ɁA
�@�W�������A���͂قƂ�Lj��܂��A�҂�S���Ȃǂ���������ɂ����Ȃ��Ă���A�̓��̗]���Ȑ������O�ɏo�������͂Ȃ�ׂ��O�ɏo���Ȃ��悤�ɁA�Z�x�̂��������A���ʂɔr�o���đ̂����Ԃ���ɂȂ�Ȃ��悤�ɂ��Ă���B�܂��A�҂̉��ލזE���牖����̓��Ɏ�荞��ł���B
�@����A�C��������ɑ�ʂ̊C��������Ő�����₢�A�̂̒��ŗ]���ɂȂ������ނ́A��Ƃ����҂ɂ��鉖�ލזE�i�������r�o�זE�j��ʂ��đ̊O�ɔr�o���A�A�͔Z�x�̔��ɔZ�����̂����ʔr�o���āA�̂̐������O�ɟ��݂����ĒE����ԂɂȂ�Ȃ��悤�ɖh���ł���B
�@�i�T����G�C�Ȃǂ̓���͑̓��ɔA�f�����߂ĐZ���������Ă���B���̂��ߑN�x��������ƔA�f���������A�����j�A�ɂȂ邽�߂ɓ��ɏL�݂�����B�j
�@��V���́A�D����ŐZ�����̒����@�\���ւ��āA��ƊC���s�������Ă����̂ł���B
�P�D�͌�����C��
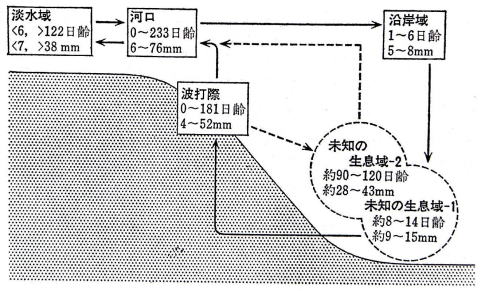 |
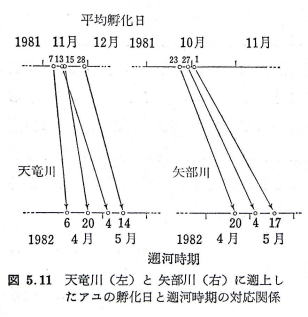
| �}�P�T�@�C�ɂ�����A���̉�V�o�H�̖͎��} �e������łƂ��A���̓���Ƒ̒��͈̔͂������� �i1988�˖{���j |
�@�����܂�̎e�A���́A�u���m�̐�����|1�v���o�R���āA���������ɔg�ł��ۂɌ����B
�@�x���܂�̎e�A���́A�u���m�̐�����|1�v���o�R���āA�x���Ȃ��Ă���g�ł��ۂɌ����B
�@�����܂�Ő����̗ǂ��A���́A�g�ł��ۂ���u���m�̐�����|�Q�v�ֈړ����傫���Ȃ��ĉ͌��Ɍ����B
�@�x���܂�Ő����̒x���A���́A�u���m�̐�����|�Q�v�ւ͍s�����ɔg�ł��ۂɑ傫���Ȃ�܂ŗ��܂�A�g�ł��ۂ��璼�ډ͌��Ɉړ�����B
�@�͂��߂̉�V�����ł́A�W�`�P�S����̎e�A���Ƒ����܂�̂X�O�`�P�Q�O����̃V���X�A���̕ߊl���ł����A���̐����悪�m�F�ł��Ȃ����B�e�A���͗V�j�͂�����قǂȂ��̂ŁA���m�F�̓���̎e�A���̐�����͋߂��̒�w���Ɛ��肳�ꂽ�̂��A���m�̐�����-�P�A�Q�ł���B
�@���̌�̒����ŁA���̖��m�F����������̎e�t�A�����\�z������w�Ő��͏��Ȃ����߂ꂽ�B
�i���m�̐�����-�P�j�́A���݈�̊C�꒼��0�`3���̒�w�ƍl�����A�z����Q�T�Ԗڂɂ����ֈڂ�Ɨ\�z�B
�i���m�̐�����-2�j�́A�g�ōۂ��킸���������i���[3�`20���j�̒�w�Ɨ\�z�B
�R���N�̒����ŁA�C�ɂ����Ă������̐�������ړ�����ۂɂ���т��āA
�@(1)�����܂�قǎ��ő����Ɉړ�����
�@(2)�����̂悢���̂قǏ��T�C�Y�ő����Ɉړ�����
���Ƃ��킩�����B
�x�R�������e���̐����ꏊ����
�i���m�̐�����-�P�j�́A�x�R�����Y������̒����D�u�͂���v�ɂ��V�N�Ԃɂ킽��x�R�p�ł̒����ɂ���āA�A���̎e���̑啔���͐��[�P�����������w�i����Ԃ�₭�������������}���ɕω�����Ƃ���j�̕\�w���i�W���̉e����������Ƃ���j�ɐ������A�\�w����Ȃ����ɏo���������Ƃ����E�ŏ��߂āi�����Ƃ��A���͓��{���ӂɂ������Ȃ����j���炩�ɂȂ����B![]() �x�R�����u�x�R�p���Ȋw����v�u�A���i�S�j�v
�x�R�����u�x�R�p���Ȋw����v�u�A���i�S�j�v
�x�R�����A�c�q�וF���̕B
�@�x�R�p�����̍��l�C�݂̍Ӕg�тł́C�A���e���� 10�`1 ���i������ 11 ���j�ɏo�����C���̕��ϕW���̒���
10 ���� 12.1�}1.8 mm, 11 ���� 18.3�}3.0 mm, 12 ���� 22.0�}4.9 mm �����
1 ���� 23.3�}2.7 mm �ł������B�Ӕg�т̉����ɗאڂ��鐅�[ 4 m �Ȑ�̐�C��ł͕��ϕW���̒�
36.1�}3.8 mm �̑�^�e���� 1�`2 ���ɍ̏W���ꂽ�B�����ώ@�ł� 11�`3 ���ɂ����čӔg�т���т���ɗאڂ����C��ɂ����Ďe���̌Q�ꂪ�m�F���ꂽ�B�x�R�p�ł�
10�`12 ���܂ōӔg�т𒆐S�ɐ������Ă����A���e���́C�e���̐����␅���̒ቺ�Ȃǂɔ����C2
�����܂łɂ͂��̉����ɗאڂ����C��֎�Ȑ�������ڂ����̂ƍl����ꂽ�B
�i�������C68(2), 144-150 (2002)�j
�@![]() �@���x�R�p�̉͌��您��т��̗אڊC��\�w�ɂ�����A���e���̏o���E���z
�@���x�R�p�̉͌��您��т��̗אڊC��\�w�ɂ�����A���e���̏o���E���z
�@�@�@�@�@�@���x�R�p�̍��l��Ӕg�ю��ӂɂ�����A���e���̏o��,�̒����z�Ɛ����ꏊ�̕ω�
�@�@�@�@�@�@���x�R�p�̘p�����Ő��炵���A���t���̉͐�ւ̉�V�k��
��04.05.07�����V��������![]() chiayu-sagami-kakou.PDF �ւ̃����N
chiayu-sagami-kakou.PDF �ւ̃����N
�_�ސ쌧���Y���������������ʎ�����ɂ��2001�`2003�N�̒����ŁA
���͐�̉͌���ŁA�A���̂Ӊ���������̎e������k����̒t���܂ł����炵�Ă��邱�Ƃ������Y���������������ʎ�����̒����ŕ��������B
�Ӊ������A���͊C�ɉ���A�����Œt���Ɉ���ĉ͐��k�シ��Ƃ���Ă������A����̒����ł͉͌����������������u�h�肩���v�̂悤�Ȑ���ł��邱�Ƃ��m�F�ł����Ƃ����B�����ɓ���������������̋Z�t���{�ւ���i�R�S�j�ɂ��ƁA�͌���ɂƂǂ܂錴���͕s�������A��̐��ʂȂǂ��e�����Ă���̂ł͂Ȃ����A�Ƃ��Ă���B
�i�l���\��̋D����ł��Ӊ���������̎e������k����̒t���܂ł����炵�Ă��邱�Ƃ��m�F����Ă��܂��B�j
�i�Q�j�V���X�A���̐����@�i�A���̐���1978���R���Y���j
�V���X�A���̉j��
�V���X�A���̉j�͂́A�����������قǑ����Ȃ�B
 �@�����Ɏg�����V���X�A���́A�l���ΕٓV����12������2���ɂ����̕߂������̂ŁA�V����R���̃A���Ɛ��肳�ꂽ�B
�@�����Ɏg�����V���X�A���́A�l���ΕٓV����12������2���ɂ����̕߂������̂ŁA�V����R���̃A���Ɛ��肳�ꂽ�B
�@�A���̑̒�4.0�`6.5�����̂��̂�p�����B
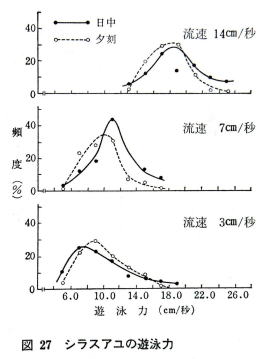
�@���H�̗����b�R�A�V�A�P�S����тT�O������4��ނŁA���������Ȃ��Ƃ�20�����j�����A���ɂ��ėV�j���x�𑪒肵���B
�@�V���X�A���͖��b14�����ȉ��̎ア�����ł͉������������̂͊F�����������A50�����̗����ł͏��`�̃A���ɗ��������̂��o���B�̒��S�������O�̃A���ɂƂ��Ă͗����T�O����/�b�͉j�͂̌��E�ɋ߂��B
�̒��ƗV�j���x�Ƃ͂قڔ�Ⴗ��B
�������A�����������Ȃ�X�s�[�h���݂�Ƃ����펯�́A����14����/�b�ȉ��ł͒ʗp���Ȃ����A�����T�O����/�b�ł͂ނ���t�̊W�ɂȂ��Ă���B
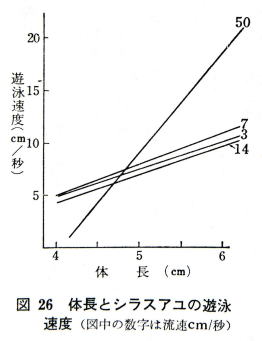
�̒�5�������O�̃V���X�A���̗����Ɖj�͂̊W���}27�ł���B
���� �R����/�b�̂Ƃ��̉j�͂́@�U�`�P�P����/�b
���� �V����/�b�̂Ƃ��̉j�͂́@�W�`�P�R����/�b
����14����/�b�̂Ƃ��̉j�͂͂P�T�`�Q�O����/�b
����14����/�b�ł͗����R����/�b�̂Ƃ��̂Q�{�ȏ�̉j�͂��������ꂽ�B
������x�̗������Ȃ��ƃA���͑����j���Ȃ��悤�ł���B
����ƐF�̊��͊C�֏o��ƕς�
�S�O�O�O lux �ȏ�ł͌���������B
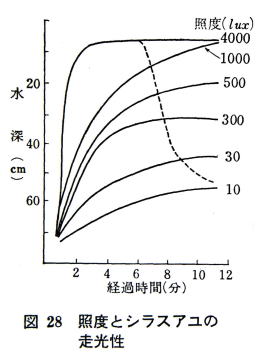
�@3.5�`5.7�����̃V���X�A���ő����������ׂ��B
�@�S�O�O�O lux �ȏ�ɂȂ�ƌ���������A���̑����������߂����B
�@�V���X�A���̑����������͂P�O lux �ł��N����A�Ɠx�̑����ƂƂ��ɂ�����ɂȂ�A�}�Q�W�̂悤�ɂP�O�O�O
lux �ł͐��ʋ߂��܂ł̂ڂ�B�������Ɠx���S�O�O�O lux �ɂȂ�ƘI���U����ɂ͈ꕔ�̃A���͉��~���Đ��[�T�O�����t�߂̑w�ɑ����W�܂�B
�@�������������畉�֕ς�̂́A
�e�A���̏ꍇ�ɂ́A1�� lux ���x�ł��������A
�V���X�A���̏ꍇ�́A��S�O�O�O lux �Ŏe�A�����ア�Ɠx�ŋt�]����B
�C���̃V���X�A���́A�����Ɠx�̕ω��ɉ����Ȃ���A���Ȃ�[���w���j���ł�����̂Ƃ݂Ȃ����B
�F���ւ̑������͒������Ⴍ�A�Ό��i520mn�j�ɑ��Ă͊��������ɒႢ�B
(4000 lux �́A�l�����̓��v�Q�`�R�O���قǑO�̖��邳�ŁA1000 lux�͓��v���̖��邳�ł���B�j
�V���X�A���͖��x���ɕq��
�i���̂P�j�̐}�X�Ɠ������u�ŁA���x���Ɣw�i�����ׂ�ƁA
�̒��R�����ȉ��ł́A����n�F�Ƃ����w�i�����A����n�F�Ƃ����w�i�ɗǂ����������B
�@�V���X�A���͔���Ȗ��邳�̈Ⴂ�����m���闧�h�Ȏ��o�������A���邢�V���w�i�ɂ��Ă���Ɖa�������Ƃ̊Ԃɐ����邲���킸���Ȗ��x�������m���ĉa����荞�ނ��̂Ǝv����B
�̒��R�����ȏ�ł́A����n�F�Ƃ����Ƃ��ɂ͏��`�̃A�������������͂邩�ɍ����B
�@����́A���x�̍����V���w�i�Ƃ������łȂ��A���x�̒Ⴂ�����⑤����w�i�Ƃ��Ă��ۉa�s������肤�邱�Ƃ��Ӗ����Ă���B
�@�C�ł̎�ȉa�͏����ȓ����v�����N�g���i�R�y�|�[�_�A�P���~�W���R�ށA�I�^�}�{���Ȃǁj�B
�@�F�ʔ������݂�ƁA�e�A���ł͗ΐF�n���̐F�ɂقƂ�ǐF�o�������Ȃ��������A�V���X�A���͗ΐF�n���̐F�ɂ�⊴���݂����S�Ă̐F�ɔ������A�e�A�����F�ʊ��o���S�ʂɍ��܂��Ă���B
�V���X�A�����ԐF��������
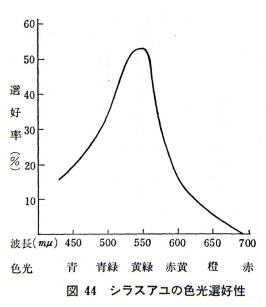
550 nm�𒆐S�Ƃ����ΐF�n�̌���������Ƃ��悭�I�D���A������g���̒���������Z��������D�܂Ȃ��B
���ɁA�ԐF�������̐����͌����ł���B
�V���X�A���͉��g���̂ǂ̐F�ʂɂ������Ă��F�o�������A�ΐF�n�F�ɂ�⊴�x���Ⴍ�A�ԐF�n�F��F�n�F�ɕq���ł���B
�ԐF����F���̓V���X�A���ɂƂ��Ď��o�h���̋�������ł���A���ɐԐF�͎��o�h���������������������A���͂���������Ɛ��肳���B
���ΐF���́A���o�h���̎ア���Ȃ̂ŁA�h���̏��Ȃ����Ό��ɏW�܂�B
�������̊ώ@�ł́A���ΐF���тɂ���A���͉����u�������v���Ă���A�ԐF���т��j���A���͂˂Ɂu���炢��v���Ă����Ƃ����B
�A�����^�ԃo�P�c�����F���ΐF���悢�B�ԐF�͑匙���B���ǂ��Ȃ��B
�V���X�A�����o�P�c�ʼn^������Ƃ��ɁA�ǂ̐F���悢�����������B
�Ԃ�̃o�P�c�̓A�����呛�����ĂȂ��Ȃ��Â܂�Ȃ��B
�≩�F�̃o�P�c�̃��A�͊��������Ɉ��肵�Ă��āA�����̓o�P�c�̖��邢���ɒ�ʂ���B
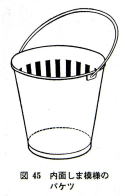 �o�P�c�̓����ɐ}�̂悤�ɍ��̃r�j�[���e�[�v��\��t���Ԋu�Q�����̏c���ܖ͗l�����A����ɃA������ꂽ�B
�o�P�c�̓����ɐ}�̂悤�ɍ��̃r�j�[���e�[�v��\��t���Ԋu�Q�����̏c���ܖ͗l�����A����ɃA������ꂽ�B
�ԂƐ̃o�P�c�̃A���͂��Ȃ范�����A�������s�K���Ɋ�������B
���Ɨ̃o�P�c�̃A���́A�ŏ������܈ȊO�̖��邢�����ɓ������A�₪�ăo�P�c�̎��͂�V�j���n�߂�B�o�P�c���Ԃ炳���A���U���ĉE�ɍ��ɉ�]������ƁA�����������ʂ͍��܂�B
�A���͔w�i�ړ��Ɛ��̓����ɓ������Đ��������݂̂ɗV�j���A��ђ��˂���̂͂Ȃ��Ȃ����B
�V���X�A���͐����ƂƂ��ɗ␅���D��
 |
 |
�@�z����������̎e�A����19�`20���̐������D�B
�@�̒��R�`�S�����̃V���X�A���̑I�D���x�̃s�[�N�͂P�U��
�@�@�@�@�@�@�P�S�`�P�V���̂Ƃ�����V�Q���̃A�����I�D�����B
�@�̒��S�`�T�����̃V���X�A���̑I�D���x�̃s�[�N�͂P�T��
�@�@�@�@�@�@�P�R�`�P�W���̂Ƃ�����W�Q���̃A�����I�D�����B
�@�̒��T�`�U�����̃V���X�A���̑I�D���x�̃s�[�N�͂P�S�D�T��
�@�@�@�@�@�@�P�Q�`�P�T���̂Ƃ�����V�V���̃A�����I�D�����B
�@�̒��U�`�V�����̃V���X�A���̑I�D���x�̃s�[�N�͂P�R�D�T��
�@�@�@�@�@�@�P�P�`�P�T���̂Ƃ�����X�O���̃A�����I�D�����B
�V���X�A���͐�������ɂ�ĐϋɓI�ɗ␅��I�Ԑ���������B
�P�����������邲�ƂɂP���Ⴂ������I�D����B
�@���c�ȉ��݈�����܂���Ă����e�t�A���͂Q�`�R���̍��������ɉ͌��ɐڋ߂��Ă���B���t�̉͐��͊C�����₽���B�ɂ�������炸�A�A�����͐��̉e��������߂Ă���̂́A���́u�␅��I�Ԑ����v�̔��B���d�v�ȗv���ƂȂ��Ă���悤�Ɏv����B
�i�R�j�A���̎��͂������A���ς�H
�@�A���́A�C�ł̓������v�����N�g�������ł̕t�����ނւƐH�����ω�����B���̐H���̕ω��ɔ����āA�̒�50�`70�����̑k������������ɂ��āA�t���^�̎��i�����Ⴍ���j���琬���^�̎��i��莕�j�֕ω�����B
�@�t���^�̎�
�@�Ƃ������~����̎��Ő����^�Ƃ͂܂������قȂ�B
�������i���傱���j�A���W�����A�������A�������A���ҍ����i�����������j����ш�������������B
�C�Y�A���͑̒���30�����A���i�ΎY�A���͑̒���35�����̍������̎����ł��n�߂�B
�̒�42�����ɂȂ鍠�ɂ͎��͏o���낢�A�̒���60�����ɂȂ邱��܂ŐL�т�B
�t���^�̎��̂����������A���W��������ю������́A�k����ɒE������B
�C�Y�A���͑̒�50�����ŒE�����͂���60�����ɒB����܂łɂ��ׂĒE������B
�ΎY�A����60�`64�����ɒB�������ɒE�����͂��߁A��������78�����A���W������72�����A��������80�����ɒB�����Ƃ��ɒE������������B
�@�����^�̎�
 �@�����^�̋��i�����傤���j�̌����́A�̒�25�`30�����̍��Ɋ{���̊O���ɂł��Ă��邪�A���G�o�̎���̉肪�`�������̂́A�C�Y�A���ł͑̒�35�`40�����A�ΎY�A����40�`45�����̂���ł���B
�@�����^�̋��i�����傤���j�̌����́A�̒�25�`30�����̍��Ɋ{���̊O���ɂł��Ă��邪�A���G�o�̎���̉肪�`�������̂́A�C�Y�A���ł͑̒�35�`40�����A�ΎY�A����40�`45�����̂���ł���B
�����^�̎��͏�{�A���{�Ƃ�12�`14�̎���ō\������A���ꂼ��̎����12�`20�{�̂ւ��̏�������ł��Ă���B
���{�̑������͔��i�ΎY�� 48�`57 �C�C�Y�� 53�`62 �B
�@![]() �@���A��"Ayu"�̎��Ɋւ��錤��
�@���A��"Ayu"�̎��Ɋւ��錤��
�i�S�j�V���X�i�C���V�j���Ŏe�A�������l����Ă���
�@�A�������̌������S���I�Ɋ뜜����Ă�����������A�_���E���̌��݂�͐�̍r�p�A�A���̏��Ȃ������Ȃǂɉ����ăA�������̌����̌����̈�Ƃ��ăV���X�i�C���V�j���ɂ�����V���X�A���̍��l���^���Ă���B
�@�V���X�i�C���V�j���͉��ݕ���ڂׂ̍����Ԃ��Q�ǂ̑D�ʼng���čs���Ă���B���̂Ƃ��V���X�A�����C���V�ƈꏏ�Ɋl���Ă��܂��Ƃ����̂ł���B���ۂɁA�V���X���t���L�q���Ƃ��L���^���E�C���V�̎q�Ƃ����݂��Ȃ����̖��O�����ċ��̂͂��̃A���̗c����߂��Ă������Ƃ��m���Ă���B���݉����ɖԂ��g���V���X���͉��x���������Ƃ����邪�A�����Ă݂�Ɗm���Ɏe�A�������l�����Ƃ����̂͋^�����Ȃ����Ƃł���B
�@�C���V�Ǝv���ăV���X��H�ׂĂ���Ƃ��A���̒��Ɏe�A�����������Ă��邩������Ȃ��B�Ȃ�Ƃ��Ȃ�Ȃ����̂��B
�C���V���ŋ߂͕߂�Ȃ��Ƃ����ĉ��N�ɂ��Ȃ邪�A�C���V�ƃA���̎����̂��߂ɁA���炭�̊ԃV���X�������ɂ��Ă͂ǂ����낤���B
�Q�D�C����͌���
�@��`�O���̂���V���X�A�����͌���Ɍ���͂��߂�B�̒��T�`�V�������x�ɐ������A���̂ɍ����A�������̃V���X�A������t�A���ɕϐg���Đ�ւ̂ڂ鏀�����͂��܂�B
�@��`�O���̑��t�͊C�̐�����̐����₽�������ŁA��ɏ������悤�ɃA���͐����ƂƂ��ɒᐅ�����D�ނƂ��������ɂ���Đ����̒Ⴂ�͌���ڋ߂��Ă���B
�������͌���ڋ߂���ɂ́A�̓��̃z�������≖�ލזE�̓�����ς��đ̉t�̐Z���������@�\���C���^����W���^��ւ��Ȃ���Ȃ�Ȃ��B
�@�t�A�����C���ƒW�������������D�����ɒ{�{���āA�W���ƊC���̂ǂ�����D�ނ������������ʂł́A
| �C�������{�{�� | �t�A���̍D�� | ||
| �W���D�� | �C���D�� | ||
| �C�������� �� |
�P�O�O�i�C���j�@ | �@ | �� |
| �V�T�i�D���j�@ | �� | �� | |
| �T�O�i�D���j�@ | �� | �@ | |
| �Q�T�i�D���j�@ | �� | �@ | |
| �O�i�W���j�@ | �� | �@ | |
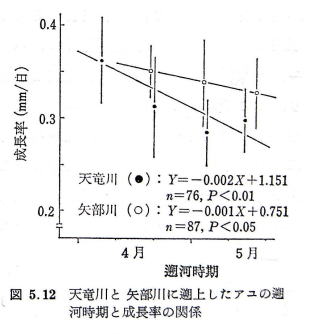 �@�������i�P�������艽�����傫���Ȃ������j�Ƒk�͎����̊W���������̂����}�ł���B
�@�������i�P�������艽�����傫���Ȃ������j�Ƒk�͎����̊W���������̂����}�ł���B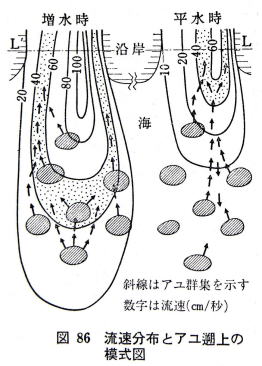
 �A�o�ʗ́A�W�����v���i�A���̐��ԁF1978���R���Y�j
�A�o�ʗ́A�W�����v���i�A���̐��ԁF1978���R���Y�j| �t�A���̏��q���x�i�����j | |
| �̒� | �œK�ȗ��� |
| 5�`6���� | 30�`50����/�b |
| 6�`8���� | 40�`60����/�b |
| 8�`9���� | 50�`70����/�b |
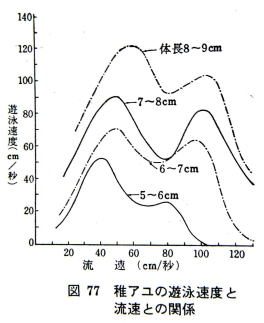
���̓K���ȗ����Ƃ��̎��̗V�j���x�́A�A���ɂƂ��Ď����V�j�s���̂��߂̍œK�ȏ����ƍl������B���ꂪ���q���x�i�����j�ł���B
�@�����������Ȃ��Ă����ƗV�j���x�͒ቺ���A�ӂ����ё��x�𑝂��đ��̃s�[�N�ɂ�����B���̃s�[�N�ȏ�̗����ł͗V�j���x�͋}���ɗ����A���ɑ����s�\�ɂȂ藬����Ă��܂��B
�V�j���x�̑��̃s�[�N�͍Ō�̗͂��ӂ肵�ڂ��ďu�ԓI�ɏo�����X�s�[�h�Ŏ������͖����B������ːi���x�i�����j�܂��͔������x�ƌĂ�ł���B
�@�A���͏��q���x�̎��Ɍp�����Ă����Ƃ������j�����Ƃ��ł���B
�����A���q���x�̗����т��̂ڂ�A���̑k��H�ƂȂ��킯�ł���B
�@�X�̃A���ɂ���ăo���c�L�����邪�A150�`200����/�b�̗����ł͒t�A���͂قƂ�Ǒk��ł����A100�`150����/�b�̗����ł���������ɋt�炦���ɗ�����Ă��܂��B
�@�̂ڂ�A���̉j�͂̌��E��120�`130����/�b�ŗ���ɋt����Ă����鎞�Ԃ͕��ς���ƂV�`�P�O�b�Ԃł���B
��ɑk�サ�Ă���t�A�������Ζʑk��͂��ǂ̒��x���邩�H
�@���R�������쒪�~���ł̒�������
�i���̎Ζʊm�x���R�O�x�A�z�����[�Q�`�R�����A�������P�T�O����/�b�j
���̎Ζʂ�o�鋗���ׂ�ƁA�̂ɂ��o���c�L���傫�����A�W�O�`�P�Q�O�����o����̂������A�ō��łP�R�O�`�P�S�O�����������B�P�S�O�������z�����̂͂P�������Ȃ������B
���Ζʂ��T�O�����ȏ�o�������̂̓o�ʃX�s�[�h�ׂ�ƁA���b�Q�T�`�R�T�����̂��̂������Ƃ����������B�܂�P���̎Ζʂ�o��̂ɂR�`�S�b��v����B
�@���̒������������R���Y�����A�u������̉��ɂ�����c��̉��ɂ���A���̎�́A�z���^�̗������ʂ͂Q���ȏ�͂���̂ŁA�t�A���̑k��͊��S�ɑj�~�����B�����h�����Ƃ���A�Ζʊp�x�����₩�ɂ��ė����������邩�A�������ʂ̒������P���ȉ��ɂ��邵���Ȃ��B���ꂪ�s�\�Ȃ�A���ʂ̓r���ɊK�i�������܂�����A�����E�X�e�b�v�����ăA�����z����Ȃǂ̕��@�����낤�B�v�Əq�ׂĂ���B
�@�i�t�A���̃W�����v�͂́A�~�����ł͕��ςQ�T�`�R�O�����A���ꂪ���萅����������ł���ꍇ�ɂ͂U�O�`�V�O�����Ƃт͂˂�B
�t�A���̃W�����v�͂��炷��T�O�������炢�̗����͔�щz����ꂻ���Ɏv���邪�A���ۂ̓W�����v�����A���������̑��Ɏ�悤�ɗ������邱�Ƃ������̂ŁA��щz�����闎���͂R�O�������炢���������B�j
�i�T�j�Ƃт͂ˍs���Ƒk�͍s���i�˖{�@���j
�@�Ƃт͂ː��̋������̂́A�悭�k�サ�A�Ȃ�萫���������B
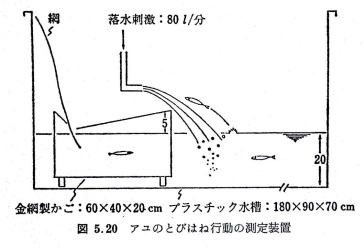 �@���}�̐����ɃA�������A�����h����^����B���̗����Ɍ������ĂƂт͂˂��A�������Ԑ������ɏW�߂āA�P�ʎ��ԓ�����̂Ƃт͂˗��i�Ƃт͂ˌ̐�/�팱�̐���100�j�����߂��B
�@���}�̐����ɃA�������A�����h����^����B���̗����Ɍ������ĂƂт͂˂��A�������Ԑ������ɏW�߂āA�P�ʎ��ԓ�����̂Ƃт͂˗��i�Ƃт͂ˌ̐�/�팱�̐���100�j�����߂��B
�l�H�͐�i����193���A��1���A���[10�`60�����A���ϗ����R�R����/�b�A�T�ӏ��͋[�̐��ƕ�����������́j�ɃA����������đk�㐫�ׂ����ʂƁA�Ƃт͂ˍs���̌��ʂƂ���v�����B
�����O�ɂƂт͂ˍs���̋���łQ�Q�ɕ������t�A�����͐�ɕ������đk�͐��ׂ��Ƃ���A�Ƃт͂ː��̋������̂́A�ア���̂��悭�k�サ���B
�܂��A�Ƃт͂ː��̋������̂قǂȂ������鐫���������A�ĕߗ����������Ƃ��m�F���ꂽ�B
���������ł́A�����㏸���ɂƂт͂ˍs���͊����ɂȂ�A���~���ɂ͗}�����ꂽ�B
��O�ɂ�����t�A���̑k�͍s���ɂ͓����ω�������B�t�A���͒��ԑk�サ�A��Ԃ͂قƂ�Ǒk��Ȃ��B���R�������ł͂Ƃт͂ˍs���ɂ������ω����F�߂��A���ԁA���ɗ[���ɂ悭�ƂԁB�[���ɂ͐������ł������Ȃ�A�}���ȏƓx�̒ቺ���N����B
�����c�Ȃ�T�C�Y�̑傫���ő̂قǂƂт͂ː��͋����B�܂��A���T�C�Y�ł���Γ���̑傫�����̂͏��������̂��悭�Ƃт͂˂�B
���R���Y���̂Ƃт͂ˊ����̊ώ@�ł́A�Ƃт͂˂͓��̏o�O�̂S���܂�����͂��܂���v�R�T����̂V���Q�O���ɏI�������B
�Ƃт͂ˊ����͌ߑO�ɒႭ�A�ߌ�ɍ����B�ߌ�̂Ƃт͂ː���70���ŁA�ߑO�̂Q�{�ɂ�����ԁB�Ɠx���}���~�����Ƃ��������U������A�Ɠx���}�㏸�����Ƃ��͊������}�����ꂽ�B�i����͌�q�̐ۉa�����̊����Ɠ����p�^�[���ł���B�j
�i�U�j�t�A���̐����i�A���̐��ԁF1978���R���Y�j
�t�A���͖�����������
��[���瑼���ցA���邢�͒������痼�[�֏Ɠx���Q������悤�ɂ��������`�̐����ɒt�A��������ƁA�A���̑����͏Ɠx�̒Ⴂ�����ɏW�܂�B
�t�A���͂Q�`�R lux �̔���ȏƓx���������m���č��Ɠx������A��Ɠx����I������B
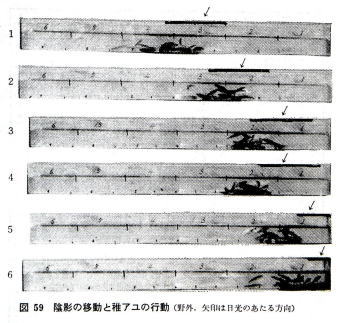
���������O�ɒu���A�����ʼne������A�P�����Ƃɍ����i�A�e�����j�����炵�Ă����ƁA���ʐ^�̂悤�ɁA�A���͉A�e�̈ړ��ƍs�������ɂ����B
�i�ʐ^��������A�ߖ��͌����̎˂������ŁA�����㕔�̍��������������e����鍕���ł���B�e�����̉��ɒt�A�����W�܂��Ă���B�j
�@�t�A�������m�ł���قǂ̏Ɠx��������A�A���͂��Ɠx�̒Ⴂ������I�����A���������֓�������o������Ȃ��B���̏ꍇ�A�Ɠx�u�����傫���قljA�e�Ŏ����͋����悤�ł���B
�@�e�A����V���X�A���̏ꍇ�ɂ͋������̑������������Ă����B�Ƃ��낪�A�t�A���ł͂��̐������t�]���ĕ����ς���Ă���B
�V���X�A������t�A���ɕϐg���邱�Ƃ��_�@�Ƃ��ĐH�̍D�݂��v�����N�g���H����t�����ސH�ɕω�����B
�v�����N�g���H�͖����w���^�Ȃ̂����A���ނւ̈Ï��w���^�łȂ���Ȃ�Ȃ��B�܂葖�����̋t�]�͐H���̕ω��Ƃ������Ȓ��a��ۂ��Ă���|�|�Ƃ������Ƃ����͒m���Ă������B
�A�e�̈ړ����x���Q�O����/�b�ȉ��ł���A���͉e�ɂ��Ă��邪�A����ȏ�̑����ł͈ړ��h���ɋ������̂��łĂ���B
�A�e�̈ړ����x���T�O����/�b���z����Ƌ����ăp�b�ƕ��U���Ă��܂��B
�i�ނ�ŁA��ʂɉe���f���ē����̂͌��ցI�����ƍ\���ē����Ȃ��B�����͖��b�Q�O�����ȉ��I�@�ށ[���Y�J�V�C�j
�ۉa�����ƌ���
�ۉa�����Ɩ��邳
�@�l�����{�̌ߌ�A�r�̒��ɃG�T��������邵�A�t�A���̐ۉa��Ԃ��ώ@����ƁA�_�̐�Ԃ��瑾�z�����˂��o���悤���Ɠx�̋}�㏸������Ƃ��͐ۉa�����͐����A�z�˂����e���ďƓx��������Ƃ��ɂ͐ۉa�����������ɂȂ��B
�A���ׂ͂̂܂��Ȃ��ɐH�������Ă���̂ł͂Ȃ��A�H�ׂ���x�肵�Ă���B�H�����Ԃ̓r���ł͏Ɠx�̏㏸�͂��܂�e���͂͂Ȃ����A�H�����Ԃ̏I��荠�ł͏Ɠx�̏㏸���������ɂȂ��āA�������Ɂu�x�ݎ��ԁv�ɓ�����̂Ɖ������B
�a��̖��邳�Ɛۉa����
�@�A���̐ۉa��͖��邷���Ă��Â����Ă��D�܂����Ȃ��B���˓����ɘI�o���ꂽ�a��ɂ͂܂������A�������t���Ȃ����Ƃ�����B����K���X�P�`�R���ŕ��������x�������Â��Ƃ���ł����Ƃ��G�T��H�ׂ��B
�a��̐F���Ɛۉa����
�@�e�a��ɓ������邳�̐ԁA���A���A�̓��ߌ�����������悤�ɂ��Ē��ׂ��B
���F���̃G�T�������Ƃ��H�ׁA�����ʼn��Ό��A�F���͂ق�̂킸���ŁA�ԐF���ł͂܂������H�ׂȂ��B
�@�t�A���͒��ˌ��̎˂��悤�Ȗ��邢�a��ɂ͂قƂ�ǂ���Ă��Ȃ��B�����Ƃ��D�܂��̂͏����Â��a��ł���B
��Ԃ≩�ΐF���̉a��͓����I�o�̉a����Â��a��ɂȂ��Ă���̂�����A���R�ۉa�ʂ������Ă悳�����Ȃ��̂����A�����͂���ɔ�����B�܂�A�t�A�����F�����ԐF���̉a���O��I�Ɋ��������B
�@�t���A�͉��F���̉a��ɂ悭�W�܂�B���Ȃ��悤�Ȑ����̓V���X�A���ł�����ꂽ���A�����͔g���̏����Z�����ΐF���т�I�D�����B
�t�A���̊������Y��
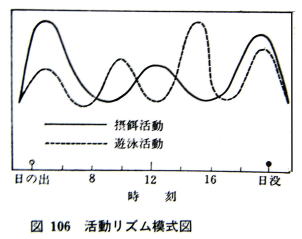
�@�Ƃт͂ˊ����͎l��^�ł���B�Ƃт͂˂͂������イ�𗣂��Ƃ����_�ŕ��ʂ̗V�j�s���Ƃ͏����Ⴄ���A�����ɋt����Ă̂ڂ�Ƃ����_�ł͂�͂�V�j�����̔��e�ɂ͂�����̂Ƃ��āA������V�j�����ƂƂт͂ˊ����ꎋ�Ă悢�ƍl����ƁA�t�A���̓����I�V�j�����͎l��^�ł���B
�@�ۉa�����́A����O��^�ŁA�s�[�N�͑����A���ߕt�߂���ї[���Ɍ����B�ۉa���Ԃ�ʂ͓����Ɠx�Ƃ������Ȃǂ̉e���ő����ϓ����邪��{�p�^�[���͌��������B
�V�j�A�ۉa�������̓����^��͎������ďd�˂����̂����}�ł���B
���̏o�O��Ɠ��v�O�͗V�j��������A�ۉa��������A�ߑO��������ߌ���̊Ԃ́A�j���ł̂��H���A�܂��j���Ƃ������ӂ��ł���B
���̂悤�Ȋ������Y���́A�̂ڂ�A����Q��A����{�B�A���ɒʗp���郋�[�����Ǝv����B
�i�̂ڂ舼�̃G�T�ނ�����Ă������̂��Ƃ��v���o���ƁA����������n�߂V�����W������ɂȂ�ƁA�s�^�b�ƐH�����~�܂�̂��ʏ�̃p�^�[���������悤�ɋL�����Ă���B���̎��Ԃ͐ۉa�����̗������ގ��Ԃƈ�v���Ă���B�Ⴂ���ɂ��ꂪ�����Ă���A�����l�o�����Ƃ��Ȃ������̂ɁA�Ǝv���B���}����A�}�S�����̃p�^�[���Ȃ̂��H�j
���o
���x���o�A���ʔw�i
�i���̂P�j�̐}�X�Ɏ������n�����w�i�ړ����u�ł̂ڂ�A���̖��x���ʔ���������ׂ��B�A���̔����͖��x�����������Ȃ�ɂ�Ⴍ�Ȃ�B
�e�A���A�V���X�A���ɔ�ׁA�ړ����x�̒x���i25����/�b�j�w�i�ɂ͂��܂蔽�����Ȃ��B
�F�ʎ��o�A�F�o
�w�i�F���������n�����w�i�ړ����u�Œ��ׂ��B
�t�A���������Ƃ���������F�́A��F�ƐԐF�ŁA���ƐԎ��ɂ͂قƂ�ǔ������Ȃ��B���A�A�͂��̒��Ԃł���B
�Ԃ��F�ɂ����Ƃ������F�o�������A���ʼn��E�A���ۂ��F�͂��܂茩�����A�����ۂ��F�͂قƂ�nj����Ȃ��|�|�g�����Z���F�قǐF�o���Ⴂ�B
�@�e�A����V���X�A���̎���́A�g���̒����ԐF�n�A���Ŕg���̒Z���F�n�ɐF�o�������A�g���̒��Ԃ̗ΐF�n�ɐF�o���ア�Ƃ�����������݂̌X���ł������B
���܂�Ă��狤�ʂ��Ă���̂́A�A���͐ԐF�n�̐F�ɂ���߂ĕq�����Ƃ������Ƃł���B
�����ȐF�A�D���ȐF
�i�ނ�l�̕����ɂ��ď������Ă���Ă���̂ŁA������Љ�悤�B�j
�A���ނ���y���ސl�����̒��ɂ́A���C�ŐԂ��̂����������̂̕��������Ă���l������B�݂��ڂɂ̓J�b�R�悳���������A�킴�ƃA�������Ȃ��悤�ɂ��Ă���悤�Ȃ��̂��B���ܕ��̏�߂͂���ɂ����Ȃ��B���Ɖ��F�̂��ܕ��ł�100���A���Ɛ��̂��ܖ͗l��80���ȏ�̃A��������������B����ƂĔ�������˂��邩��K���łȂ��B�����Ĕh��ȐF�ʂ͒ނ蒅�ɂ͍D�܂����Ȃ��Ƃ������Ƃł���B
����̐F���炢���ƁA�t�A���͉��F���D�����B
�ނ蒅�ɐԂ������Ȃ��Ȃ物�F�͂ǂ����B���F�̕��������Ă���A�����W�܂��Ă���̂��A�Ƃ����Ƃ���͂�����Ȃ��B���F�͔��ɖ��邢�F�Ō������������˂��邩��ł���B�����Ƃ��A����𑽂����˂��Ȃ��悤�Ȑ��n������Εʂ��B����ȐS�z�������莇�n���̐F���𒅂��ق�������Ƃ������̂��낤�B
�@�����Ӗ��ŁA�ނ�Ƃ��ނ莅�����F���悢���ƂɂȂ邪�A�Ƃ̂ق��͉e�Ƃ����h���������̂ŁA���قǂ̌��ʂ͊��҂ł��Ȃ��B
�����̑I�D��
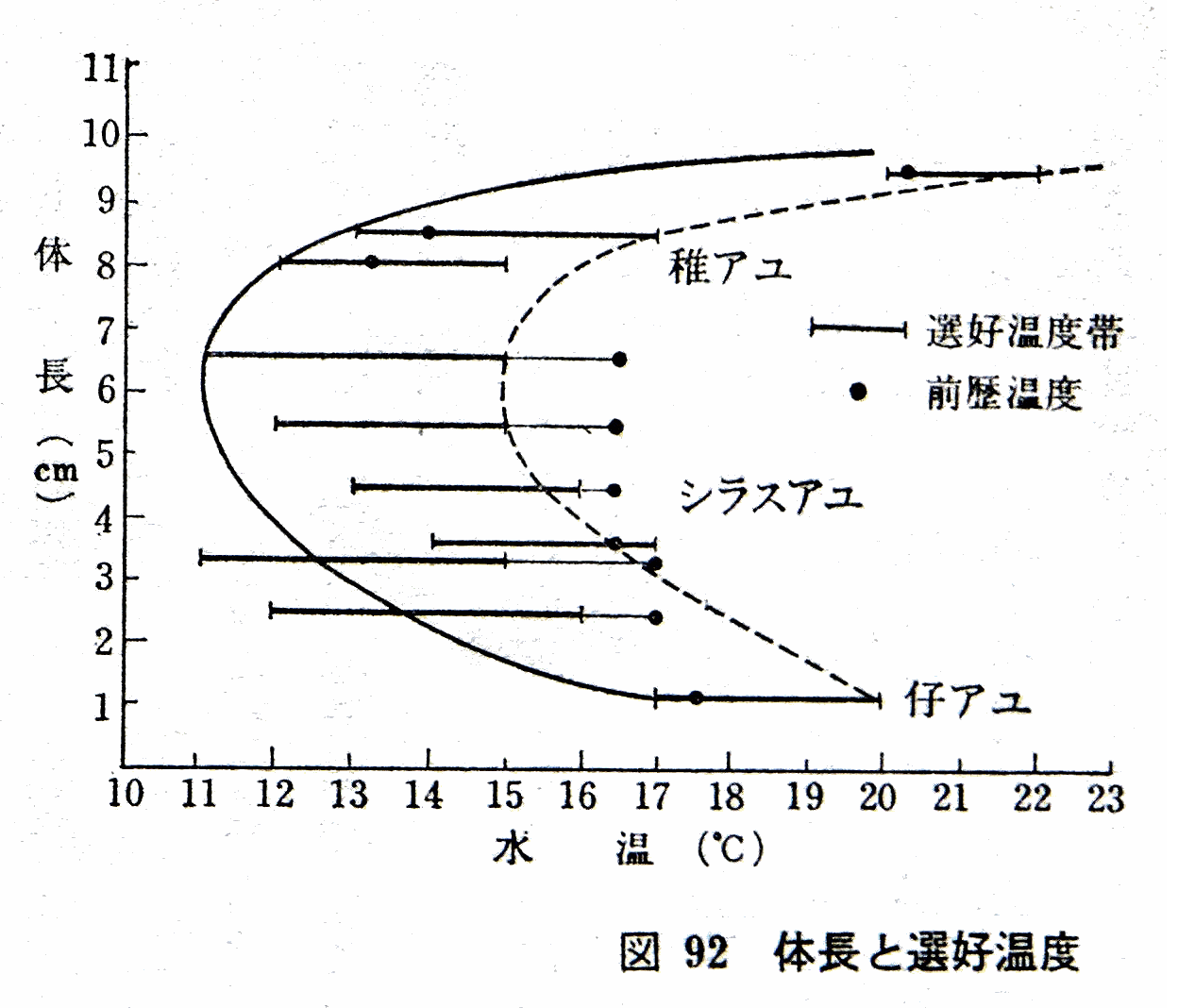 �@�e�A������t�A���܂ł̑I�D���x�̐��ڂ߂Ă݂悤�B
�@�e�A������t�A���܂ł̑I�D���x�̐��ڂ߂Ă݂悤�B
�e�A���ł͑O�����x��荂�߂̐�����I�сA
�V���X�A���ł͂ނ���ቷ��I�сA
�t�A���ł͂ӂ����э������̐�����I�ԁB
���̂��Ƃ̓A���̐����Ƃ͐��Ă���Ȃ��K���ł���B
�@���t�̉͐��͗₽���B�������A���͈�`�I�ɐ�i�W���j�ɂ̂ڂ邱�Ƃ�]�V�Ȃ������B�g�����C���ɂ���A���������O�����x��荂���������D�ނƂ���A�A���̑k��͑啝�ɒx��A����̐����Ɏx����������B�����A�K���Ȃ��ƂɁA�k��O�̃��A�͑O�����x����`�l�x���Ⴂ������I�Ԃ悤�ɔ�����Ă���B���A�͉͌��[�����肱�݁A�������ɑ̒��𐮂��đk��̋@���҂B�₪�ĉ͐����x���O�����x�ɋ߂��Ȃ�ƁA�k����J�n����B
���̈Ӗ��Łu�C�������͐����ƂقڂЂƂ����Ȃ�Ƒk�シ��v�Ƃ����̂͐������B�������A�C�����Ƃ������A�D�����Ƃ������ق��������Ɠ������Ă���B
��ɂ̂ڂ����A���̑I�D���x�͏����������Ȃ�A�O�����x��萔�x�����Ƃ��������Ԃ��A���̏���͓��`���x���炢���Ǝv����B
�����I�D��
�@�A���͐��ݐ����D�݁A���萅���������Ƃ͈��ނ������҂͒N�ł��悭�m���Ă��邪�A�����ڂ������Ă��Ȃ��Ă����w��������p���ʼn������ꂽ���͌����邩�Ƃ����ƁA����͂��܂�m���Ă��Ȃ��B
�ؑ]�O���̒��ǐ�A�K���A�ؑ]��̐��ɑ���̂ڂ�A���̑I�D��������ׂ邽�B
�����A���͎O�d�����c��Y�ő̒��V�D�R�����Œn�����Œ{�{�������̂�p�����B
���ǐ�ƗK���̐�������ׂ��ꍇ�����ǐ�Ɩؑ]��̐�������ׂ��ꍇ���A�A���͒��ǐ�̐����D�B
�M�Z�����������̓�[�ɂ������������ŁA�V���s���^�������邱�Ƃ���ړI�ɂ��āA��͒Õ����H������A����Ɋ֒J�����H�����ꂽ�B���Ƃ��Ƃ̐M�Z��͌����iCOD2.56ppm)�Ɗ֒J�����H�̐��iCOD1.64ppm)�A��͒Õ����H�̐�(COD0.4ppm)�Ƃ�����ׂ�ƁA�����H�̐����D�܂ꂽ�BCOD�̍��i���̉���j�����̂܂ܑI�D���ɔ��f���Ă���B
�]�̐�̃p���v�H��p���ł̎����ł́A�H��p�����ł̔r�����t��COD��2340�������ł������B���̔r�����t���ł͂W����ɂ̓A�����S�Ď��B���{�܂Ŕ��߂���������Ȃ��Ȃ邩��������ƁA�p�t���P���{�Ɋ�߂��Ă悤�₭�͐��Ƌ�ʂ��Ȃ��Ȃ����B���̂Ƃ���COD��0.23�������ł���B
�i�c�O�Ȃ���A���̂Ƃ��p�t���̂ǂ̐������A�������������̂��͒��ׂ��Ȃ������B�j
�@��L�̐M�Z��A�]�̐�̗�ł��킩��悤�ɁA�쐅�̉���͊ԈႢ�Ȃ��A���̑k���j�Q���Ă���B
�i�V�j�t�A���́A�P�O�����̈�Ƃ����������ʂł�������܂̏L�����������Ĕ�����B
�@�u�C�ɂȂ�b�v���g�k�`�r�Ƃ͂ǂ�Ȃ��̂��h�ɂ����������A�����搶�ɂ��u���ނɂ�鉻�w��܂̊��������@ �\�A���ɂ���܂̊���
(1978)�v�ł́A�����E�ʊ����܂k�`�r�͂킸���P�D�T�������ł��A���͂��̏L�����������邱�Ƃ��m�F����Ă���B�Q�ƁG������Y�́u���I�@�ނ�Â�̋L�v�@��ǂ���B
�@�\�N�ȏ���O����A���̑k�オ�������ƌ���ꑱ���Ă���B
�@���̌����̈���A�ƒ�ȂǂŎg��ꂽ������܂��͐�ɗ���A���̏L�����������Ēt�A�����͌��ɋ߂Â��Ȃ��Ƃ����̂��B
������
�@�������Ƃ́i�P�O�O�����̈�j�P�O�����̕����P�O�O�O���b�g���̐��ɗn�������Ƃ��̔Z�x�ł��B
�@�������Ƃ͂������̐番�̈�̔Z�x�i�P�O�����̈�j�ŁA�P�O�����̕����P�O�O�O�g���̐��ɗn�������Ƃ��̔Z�x�ł��B
�@��~�ʂ̏d�����P�O�����ł��B
�@����@�Ő���Ƃ��ɁA��Q�O���̐�܂��g���܂��B�Q�O���̍�����܂��P�������̔Z�x�ɔ��܂�ɂ͂Ȃ�ƂQ���g���̂��ꂢ�Ȑ����K�v�ł��B����������ɗ����ƁA�P����邽�тɁA���ꂢ�Ȑ��Q���g���Ŕ��߂Ȃ���A�A���͂k�`�r�̏L�����������đk�サ�Ă��܂���B�i�������D�����̑�^�r��������ȊO�ł͂k�`�r�Ȃǂ̍�����܂͏����ł��܂���B�܂��A��ɗ��ꂽ������܂́A��ŕ�������Ȃ��܂܊C�ɏo�ĉ͌��߂��̊C��ɒ��a���Ă��邻���ł��B�j
�@���t�́A������܂��g��Ȃ��ʼn������B�̂���̐Ό��ŏ[�����ꂢ�ɐ��ł��܂��B
�@�t�̂̐���p�Ό����̔�����Ă���A�䂪�ƂŎg���Ă��܂��B����ō�����܂Ɠ����ɐ���ł��܂��B
�@�܂��A������܂ɓY������Ă���u�������܂́A�H�i�q���@�A�@�ŐH��A�z�ЁA�����p�悾�ꂩ���E�����E���ނA��сA�K�[�[���ւ̎g�p���֎~����Ă��܂��B�^�������Ȃ�Ɛ�`����Ă��鍇����܂́A�l�ɂ��A���ɂ��댯�ȏ��i�ł��B
������
�k�㎞�̏�Q�Ɗ댯
�@�悤�₭�t�A�����͌������Q��ƂȂ��đk����͂��߂Ă��A�C�̖L�x�Ȓ�����ɓ�������܂łɂ͑����̊댯�Ə�Q������B
�@�ő�̓�ւ́A���A����H��_���ƁA�t�A���̑k��Ȃ��`�������̋����Ƃ������O���������H�ł���B
���⓪��H��_���̉��̐[�݂ɏW�܂����t�A���̓J���E�ɍU������A�̂ڂ������̘e�ł̓T�M���҂�������B���⓪��H��_���́A�ނ�Ɋi�D�̉a�����Ă���̂��B
�@�ߔN�J���E���ُ�ɑ����Đ��ł̔�Q���}�����Ă��邪�A���̈���͐l�ɂ����j��ɂ���Đ��Ԍn�̃o�����X�����ȕ����֕���Ă����Ă��邩��ł͂Ȃ��낤���B
�@�i�W�j�������x�̑����t�A�����Ȃ��`�����ł�����
�@�@![]() �@�������x���قȂ�A����c�̂Ȃ�萫
�@�������x���قȂ�A����c�̂Ȃ�萫